研究内容
2017
神経系の複雑な機能は神経回路の中での情報処理によって行われています。シナプスからの情報の入力、情報の統合、迅速かつ正確な情報の出力が神経回路の機能維持のためには重要です。近年、この神経回路の形成や機能にグリアが重要な役割を果たすことが明らかになってきました。しかし、その詳細なメカニズムに関しては、まだ不明な点が数多く残されています。また、ヒトの神経難病の病態発生機序や治療法の開発に関しても、明らかにすべき点が多くあります。機能形態学教室では、特に神経細胞の出力系である軸索機能に対するグリアの役割に着目して研究を進めています。
機能形態学教室で行っている主な研究は
下記のとおりです。
1)ミエリン形成グリアによる軸索機能調節に関する研究
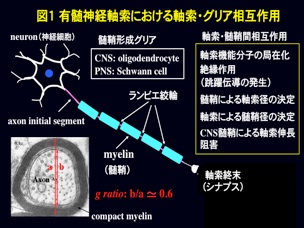
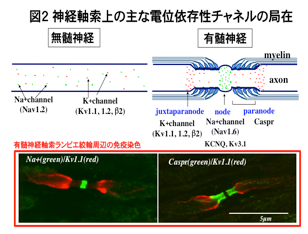
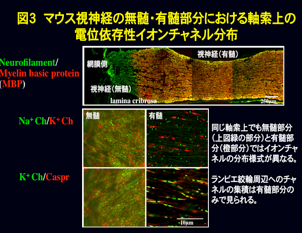
有髄神経軸索表面を覆うミエリン膜は、中枢神経系ではオリゴデンドロサイト、末梢神経系ではシュワン細胞によって作られます(図1)。一般的には絶縁膜として跳躍伝導にはたらくことが知られていますが、実はこの膜が形成されることにより、軸索は大きく変わります。たとえば軸索の活動電位発生に関わる電位依存性イオンチャネルは、無髄神経軸索では軸索表面に均一に分布していますが、ミエリン膜が形成されると電位依存性ナトリウムイオンチャネルはランビエ絞輪、カリウムイオンチャネルはジャクスタパラノード部分に特徴的に集積します(図2、図3)。この集積が跳躍伝導に重要なはたらきをしているのです。この他にミエリンあるいはミエリン形成グリアは軸索径や軸索内輸送にも関わることが知られています。私たちは、ミエリンがどのようにして軸索に影響を及ぼしているのかを研究しています。現在、ミエリン異常に伴って小脳プルキンエ細胞軸索や末梢神経軸索に生じる異常のメカニズムなどを調べています。
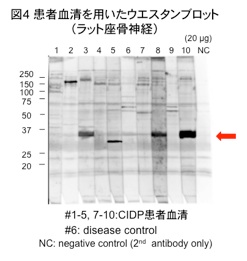
2)末梢神経障害患者血清中の抗神経抗体に関する研究
末梢神経障害を呈する代表的な疾患の一つとして免疫性
ニューロパチーがあります。これは、急性に経過するギラン・
バレー症候群とその亜型であるフィッシャー症候群、慢性
に経過する慢性炎症性脱髄性ニューロパチー(CIDP)に大別
されます。いずれも本来身体を守るはずの免疫系が神経を
攻撃することによって神経障害がおきると考えられています。
この中で軸索が障害されるタイプのギラン・バレー症候群や
フィッシャー症候群では、血清中の抗ガングリオシド抗体が
病態発生に関わります(ガングリオシドは神経系に多い糖脂
質の一種)。しかし、ミエリンが破壊されてしまう脱髄型は何故
おきるのかまだわかっていません。そこで、私たちは岐阜大
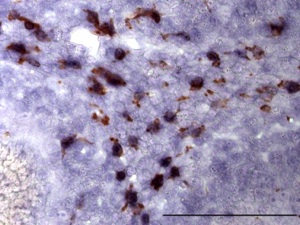
学医学部や九州大学医学部、国立病院機構宇多野病院、同国府台病院、鎌ヶ谷総合病院などの病院との連携により、患者血清中の抗神経抗体の有無、抗体と病態との関連性を調べています。現在、特に慢性炎症性脱髄性ニューロパチー患者血清中にみられる抗体の一つに着目して研究を進めています(図4)。私たちはこの抗原分子を新たに同定し、非常に抗原性の高いミエリンタンパク質であることをつきとめました。この分子自体の機能および病態との関連性に関して現在研究中です。
3)末梢神経における脱髄保護分子の解析
アネキシンIIはリン脂質および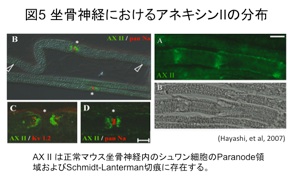 カルシウム結合タンパク質で、様々な細胞機能に関与しています。末梢神経ミエリンでは、パラノードやシュミットランターマン切痕に局在します(図5)。
カルシウム結合タンパク質で、様々な細胞機能に関与しています。末梢神経ミエリンでは、パラノードやシュミットランターマン切痕に局在します(図5)。
(Hayashi et al, 2007)。パラノード異常を示すマウスではアネキシンIIが増えますが、その役割はまだわかっていません。私たちは、リゾレシチンをマウスの坐骨神経内に投与して作製したリゾレシチン誘導脱髄モデルマウスを用いてアネキシンIIの役割を調べてきました。アネキシンII特異的SiRNAを用いた実験などから、アネキシンIIが脱髄巣の拡大に対して抑制的に関与することがわかりました。現在、この分子メカニズムに関して研究中です。
4)活性化ミクログリアに発現するPLD4の機能に関する研究
PLD4はPLD1やPLD2と異なり膜貫通型の新型PLDファミリータンパク質です。小脳で発達段階特異的に白質の細胞に発現する分子として理化学研究所古市貞一先生によって見出されました。PLD1、PLD2と異なりいわゆるPLD酵素活性を示さないことから、その機能はこれまでにわかっていません。私たちは、古市氏との共同研究で、PLD4が小脳発達段階で白質に一過性に出現する活性化ミクログリアに特異的に発現することを見出しました(図6 Yoshikawa et al, 2010)。また、発達段階のみでなく、脱髄病態で活性化するミクログリアにも存在することを明らかにしました。現在、培養細胞およびノックアウトマウスを用いてPLD4の機能を解析中です。これらは古市先生をはじめ、自然科学研究機構生理学研究所池中一裕教授(脱髄マウス提供)、農業生物資源研究所木谷裕先生(ミクログリア細胞株提供)、新潟大学脳研究所﨑村建司教授(ノックアウトマウス作製)との共同研究によって行っています。
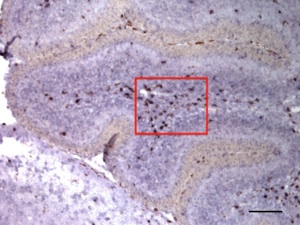
図6 この図は生後7日目のマウスの小脳のPLD4 mRNAとミクログリアのマーカーであるIba1との二重染色
下図は上図の赤枠の拡大 Bar=100μm